Familie CHRYSOMELIDAE
Subfamilie Chrysomelinae
Lacordaire, 1845
Goudhaantjes
Maak een keuze uit de begrippen of de kaarten,
of klik op TABEL hiernaast.
|
Familie CHRYSOMELIDAE
Subfamilie Chrysomelinae Lacordaire, 1845 Goudhaantjes |
|
|
|
|
|
Maak een keuze uit de begrippen of de kaarten, of klik op TABEL hiernaast.
|
Biogeografie
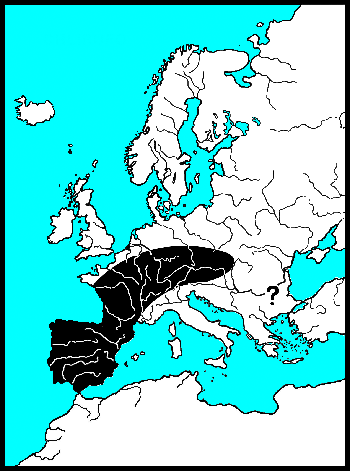
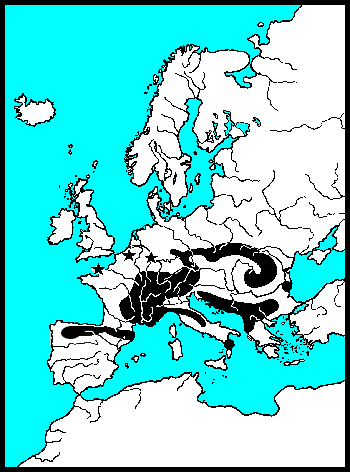
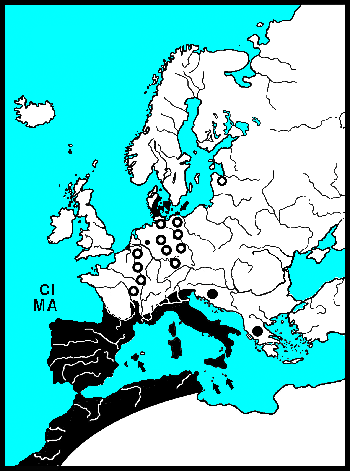
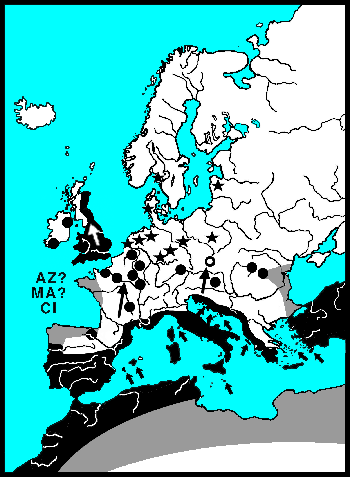
De waardplanten van wilgenhaantjes komen vooral voor in het stroomdal van rivieren en staan vaak periodiek in het water. Wilgenhaantjes kunnen ten opzichte van andere goudhaantjes goed vliegen. Het voorkomen van de verschillende soorten wordt vooral bepaald door het stadium waarin een droog habitat nodig is. Griendhaantjes (Phratora-sp.) overwinteren in holtes van de stam, zodat 's winters en in het voorjaar de bomen onder water kunnen staan. Zij volbrengen echter hun popstadium in de bodem onder de boom. In de zomer moeten de bomen waarop zij leven dus droog komen te staan. Het Veelkleurig wilgenhaantje (Plagiodera versicolora) en Chrysomela-soorten kleven hun pop aan het blad. Zij kunnen bomen op hun menu zetten, die zomers ook onder water staan. 's Winters moeten de Chrysomela's echter naar een hoger gelegen landschap vliegen, want overwinteren doen zij onder bladeren in droge bosbodem. Het kleine monofage Veelkleurig wilgenhaantje heeft drie tot vier generaties per jaar en kan zijn aanpassingen waarschijnlijk daardoor snel doorgeven. Voor hun overwintering zijn zij veel minder kieskeurig en vallen gewoon in de herfst met de bladeren mee. De roodbruine wilgenhanen van het geslacht Gonioctena moeten voor zowel hun pop als voor hun overwintering naar droge bodem. Zij leven meer in droge bossen en verspreiden zich over het algemeen langzaam. Goudhaan-soorten (Chrysolina-sp.) leven op kruiden en vliegen praktisch nooit, zij lopen vooral langs lijnvormige landschapselementen. De meeste Nederlandse dieren hebben weliswaar volledige vleugels, maar hun vliegspieren zijn gereduceerd. In Nederland zijn alleen Chrysolina graminis en C. americana vliegend waargenomen, Bourdonné geeft in Winkelman & Debreuil (2008) veel waarnemingen uit Frankrijk.
De Coloradokever heeft zich 20e eeuw met gemiddeld 60 km per jaar naar het noorden uitgebreid. Deze grote exotische soort breidt zich vooral vliegend uit. Van onze inheemse goudhaantjes vliegen de soorten van het geslacht Chrysomela het beste. Bij de recente invasiegolf van Chrysomela lapponica, die Beenen (1985) beschrijft, komt de uitbreiding ongeveer uit op 10 km per jaar. In 1990 vond S.C. Langeveld hem zelfs al in de Amsterdamse Waterleidingduinen. De lopende goudhanen (Chrysolina-sp.) komen niet verder dan 100 meter per jaar. Samen met de vleugelloze Reuzenhanen (Timarchinae) zijn goudhanen de slechtste verspreiders, die vooral de zoomen van landschappen bewonen. Het betreft hier ware olifanten op schaal 1:500, met rudimenten van vleugels. En voor deze lopende dieren gaat passieve verspreiding, meegevoerd worden met rivierwater, vele malen sneller.
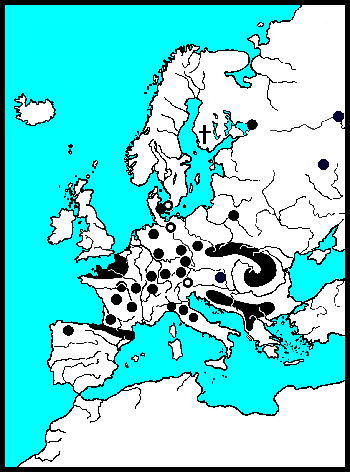
In de laatste IJstijd is Noordwest-Europa bijna geheel bedekt geweest met ijs. Europa kende beperkte uitwijkmogelijkheden in de laatste IJstijd. Gematigde soorten werden tot ver achter de ijsgrens gedreven, naar het land langs de Middellandse-Zee. Echte warmteminnende soorten naar Noord-Afrika. Veel soorten waren door de koude/droogte geïsoleerd geraakt. Door deze gebeurtenissen, als IJstijden, destructie van landschappen door de mens, overstroming, verdwaling e.d., raken groepen kevers geïsoleerd van hun verwanten. Hun verwanten kunnen zich anders ontwikkelen. Soms veranderen zij zodanig dat zij niet meer met elkaar kunnen reproduceren. Sommige soorten veranderen sneller dan anderen. Timarcha ken al separatie op het Kanaal-Eiland Chaussy dat pas sinds 1200 een eiland is (Bechyně 1950).
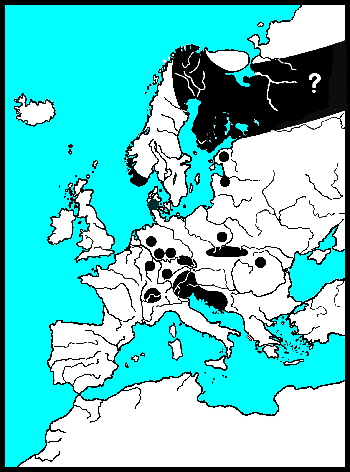
Laatglaciaal luiden vooral de pioniers onder de Composieten het einde van de IJstijd in (Van Huissteden & Ran 1993). Ongeveer 13.000 jaar geleden is er een toename van Alsem (Artemisia). Alsem is de waardplant van Chrysolina (Chalcoidea) en mogelijk ook van Chrysolina (Craspeda), deze zijn alle donker van kleur en roodgerand. Overblijfselen van deze levensgemeenschap zijn zelden, en nooit oorspronkelijk. De Berk en de Ratelpopulier waren de eerste bomen. Voor meer soorten van verschillende genera geldt dat de soorten die in het Arctisch gebied voorkomen, als laatglaciaal fossiel bekend zijn (Lemdahl & Gustavson 1997). Zij zijn vaak scherp en opvallend getekend, bijvoorbeeld Hydrothassa hannoveriana en P. phelandrii.
In veel genera vicariëeren soorten in het Palearctische en Nearctische region, en slechts enkele worden als één soort beschouwt met een holarctische verspreiding. Hun huidige status is taxonomisch weergegeven naar de laatste inzichten. Gastrophysa polygoni en meerdere Phaedon-soorten zijn echte holarctische soorten. Volgens (Lindroth 1957) is Chrysolina staphylaea transatlantisch verspreidt, over de landbrug van Europa (Schotland) via IJsland naar Amerika. Deze landburg zou tot in het Tertiaire Plioceen en het Quartaire Pleistoceen hebben voortgeduurd.
Daarnaast zijn er vicariëerende soorten, soorten die opgesplitst zijn door geografische scheiding: Chrysomela saliceti is verwant met de Nearctische C. crotchi Brown, 1956; Chrysomela heeft een duidelijkere vicariantie in het Arctische gebied, dan Gonioctena. Phratora polaris met P. hudsonia Brown, 1951 en P. frosti Brown, 1951. En verder de recent door de mens geïmporteerde soorten als de schadelijke Plagiodera versicolora en biologische bestrijders uit Chrysolina.
In
Nieuw
Zeeland
en
Australië
(PAC,
AUS
en
NZL)
40
genera
met
veel
endemische
Chrysomelinae
(vooral
tribus
Paropsini).
Zij
komen
vooral
voor
op
jonge
planten
van
de
tropische
Myrtaceae
en
Vlinderbloemen
(Fabaceae).
De
kenmerkende
Paropsis
Olivier
1807
op
jonge
Eucalyptus
L'Héritier
de
Brutelle
(Myrtaceae)
en
Calomela
Hope,
1840
op
Acacia
Miller
(Fabaceae)
(Selman,
1979) (zie ![]() literatuur). Zij
vertonen
waarschijnlijk
een
functionele
monofagie,
want
hun
verwanten
in
het
Oriëntaalse
region
eten
Rosaceae
(Jolivet
&
Hawkeswood,
1995). Ook
komen
ze
voor
in
Nieuw-Zeeland
dat
veel
op
de
Australische
fauna
lijkt.
literatuur). Zij
vertonen
waarschijnlijk
een
functionele
monofagie,
want
hun
verwanten
in
het
Oriëntaalse
region
eten
Rosaceae
(Jolivet
&
Hawkeswood,
1995). Ook
komen
ze
voor
in
Nieuw-Zeeland
dat
veel
op
de
Australische
fauna
lijkt.
In het Pacifische region komen zij niet voor op de oceanische eilanden, maar uitsluitend op continentale eilanden: de Fidzji-Eilanden en West Samoa (Gressitt 1957). In het Antarctische region 2 genera op Tierra del Fuego, de Magellan Archipel en Patagonië. Deze vertonen overeenkomsten met Australische en Afrikaanse genera. Een hiermee verwant genus in Australië leeft op de Antarctische beuk (Nothofagus). Deze bomen zijn een Tertiairrelict, zij komen zowel in Australië als in Zuid-Amerika voor. Het voorkomen van verwante Chrysomelinae in Australië, Zuid-Amerika en Afrika duidt op een ontstaan van de onderfamilie in de Midden-Jura toen Gondwanaland nog bestond.
Oriëntaalse fauna (ORI) is weinig vertegenwoordigd met 4 genera en zonder endemische genera. Vooral Phyllocharis Dalman, 1824 op de planten uit de IJzerhardfamilie (Verbenaceae) en Chalcolampra Blanchard, 1853 op Helmkruiden (Scrophulariaceae) en Lipbloemigen (Lamiaceae).
In Noord-Amerika (NEA) geen endemische genera. In totaal 15 genera, 130 soorten, waarvan de oorspronkelijk Neotropische Calligrapha Chevrolat in Dejean, 1836 het grootste genus is met 38 soorten. Zij komen nog in Zuid-Amerika voor, vliegen goed en eten specifieke struiken en bomen van allerlei families.
Zuid-Amerika (NEO) heeft het grootst aantal soorten in 31 genera, waarvan Doryphora Illiger, 1807 het opvallendste genus is (zie ook Eberhard 1981)
In Afrika (AFR) zijn er slechts 19 genera, waarvan vele endemisch. In tegenstelling tot Zuid-Amerika zijn zij uit Afrika nauwelijks bekent uit het oerwoud. De bekendste zijn Chrysolina L. en Ceralces Gerstaecker, 1855. De genera van Madagaskar zijn verwant met de genera van het vaste land.
Voor een overzichtelijke bespreking van de mondiale biogeografie, zie Daccordi (1996).