Familie
CHRYSOMELIDAE
Latreille,
1802Bladkeverfamilie
Subfamilie
Chrysomelinae Lacordaire,
1845
Goudhaantjes
inleiding
Op
bijna
elke
wilde
plant
leven
bladkevers,
maar
niet
elke
kever
op
een
blad
behoort
tot
deze
familie.
Men
zal
de
kever
nader
moeten
bekijken.
Maar
het
pakken
of
vangen
is
in
eerste
instantie
af
te
raden,
omdat
dan
alleen
het
vluchtgedrag
te
bestuderen
is.
Hier
poog
ik
juist
een
aanzet
te
geven
om
de
levenswijze
te
bestuderen.
Zonder
de
levenswijze
te
kennen
blijft
het
bij
een
eenmalige
ontmoeting,
en
is
studie
of
beheer uitgesloten1.
Maar
ook
onderzoeken,
iets
veranderen,
leert
ons
veel,
mits
we
het
dier
niet
uit
het
landschap
verliezen.
Van
alle
op
bladeren
levende
insecten
vertoont
vooral
de
bouw
van
de
voet
grote
overeenkomsten.
Goudhaantjes
hebben
zes
voeten
met
vijf
leden,
maar
het
4e
voetlid
is
klein
en
verborgen
in
het
lobvormige
3e
lid.
De
voet
heeft
een
borstelige zool (zie foto waarnemingen.be)
en
geeft
de
kever
een
uitstekende
grip
op
het
gladde blad (zelfs onder water werkt dit (Hosoda
& Gorb 2012),
waarbij
een
klauw
achter
een
randje
de
voet
stevig verankerd (zie Howard
2008).
Desondanks
is
een
voorzichtige
benadering
geboden.
De
kevers
zien
de
veldbioloog
en
ontsnappen
aan
't
oog
door
hun
poten
in
te
trekken
en
van
het
blad
af
te
rollen.
Dit
is
eenvoudig
te
verhelpen
door
te
voorkomen
dat
men
een
schaduw
op
de
plant
werpt
en
eventueel
een
hand
onder
de
kever
te
houden.
Houden
zij
hun
sprieten
recht
vooruit
en
zijn
zij
ongeveer
even
lang
als
een
poot,
dan
heeft
men
met
de
bladkeverfamilie
te
doen.
Deze
familie
kent
in
Nederland
305
soorten.
Goudhaantjes
vormen
de 59
soorten
van
de
onderfamilie Chrysomelinae.
Larven
en
kevers
leven
vrij
op
bladeren
van
bloemplanten,
meestal
in
pioniervegetatie.
Diverse
strategieën
van
levenswijze,
larven
in
de
warme
of
natte
periode,
overdag
in
schermer
of
uitsluitend
in
de
nacht,
samen
of
gescheiden
van
de
volwassen
kevers.
Bij
de
meeste
soorten
overwinteren (hibernatie)
de
kevers,
maar
er
zijn
ook
larven,
als
Gastrophysa
viridula,
die
koude
beter
verdragen
dan
de
kevers
en
dan
overwinteren
zij
(Rossolimo
1991).
In
wilgenbos
zijn
de
larven
beschermd
tegen
lopende
roofdieren,
doordat
de
planten
in
het
water
staan.
Bovendien
hebben
de
larven
van
deze
dieren
afweerklieren,
welke
tegen
Loopkevers
en
Mieren
bescherming
biedt
(Cox
1996). Zie ook de foto's van www.aramel.free.fr.
Dit
geldt
ook
de
poppen
die
in
waterrijk
gebied
bijna
altijd
extern
op
het
blad
zitten.
Er
zijn
echter
ook
roofdieren
die
juist
door
de
afweerstoffen
van
de
kevers
worden
aangetrokken
(Köpf
et al.
1997). Mohagheghi (2007) geeft
een uitgebreid overzicht van de synthese. De
bekendste
roofdieren
in
wilgenbos
zijn
vliegende
insecten
van
de
wantsen (Heteroptera)
de
algemene
Anthocoris
nemorum
(Linné),
maar
vooral
soorten
van
de
Boomwantsen (Pentatomidae
- Asopinae)
als
Arma
curtus
(Fabricius)
en
Troilus
luridus
(Fabricius);
van
de
Zweefvliegen (Syrphidae)
vooral
Parasyrphus
nigritarsis
(Zetterstedt),
van
de
Sluipvliegen (Tachinidae)
Meigenia
Robineau-Desvoidy
en
Steiniella
Malloch;
en
van
de
Plooiwespen Symmorphus (Hymenoptera: Eumeninae) (Budriené
2003 en 2004); maar voor de Britse Timarcha ook Valken, Vossen en Dassen en
Uilen, een uitgebreide systematische opsomming is te vinden in (Cox
2007).
Of
Mijten (Acari)
slechts
meereizen
of
parasiteren
op
de
kevers
is
onbekend,
een
lijst
is
te
vinden
in
Cooreman
(1950).
Gaasvliegen (Chrysopidae)
Chrysoperla
carnea
(Stephens);
de
Spiegelkever
Saprinus
virescens
(Paykull),
van
de
Lieveheersbeestjes
Adalia
bipunctata
Mulsant,
Calvia
Mulsant,
Coccinella
Linné,
Hippodamia
Dejean
en
Oemopia
L. De 'haantjes' kennen een hele biodiversiteit aan belagers, zie
bijvoorbeeld (www.koleopterologie.de).
Normaal
verkrampen
de
kevers
bij
verstoring (thanatose)
en
vallen
op
de bodem (zie en vergelijk foto's Alain Doornaert waarnemingen.be
en van Paul en Marianne waarnemingen.be),
maar
juist
blijven
zitten
komt ook voor
bij
de
Struikhaantjes
Gonioctena.
Zij
plegen
broedzorg
en
wensen
de
eerste
tijd
hun
kroost
te
beschermen
tegen
mieren,
wantsen
e.d. (zie foto's Vladimir Bryukhov - Владимир
Брюхов 2014 insecta.pro-50204).
Bij een Australische soort is zelfs vastgesteld dat de kever de
larven naar elkaar toe drijft (Reid et al.
2009). Ook in Zuid-Amerika zijn bijzondere voorbeelden (Windsor
et al. 2013).
Waardplant
De
meeste
bladkevers
beperken
zich
tot
een
bepaalde
groep
planten. Van schade wordt gesproken als de kever een voor de mens
belangrijke plant remt in groei en ontwikkeling (zie bijvoorbeeld waarneming.nl-55763236
en zie
voor
een
mondiaal
overzicht Booth
et al. 1990).
Veel
Nederlandse
namen
wijzen
hierop:
aspergehaantje, elzehaantje,
zuringhaantje
enz.
Een
goede
plantenkennis
is
dus
wel
noodzakelijk
voor
het
bestuderen
van
deze
keverfamilie.
De
kevers
die
slechts
binnen
één
plantengeslacht
hun
waardplant
hebben,
worden
monofaag
genoemd.
Soms
gaat
dit
zover
dat
de
kevers
eerder
sterven
dan
een
andere
plant
eten!
Het
merendeel
eet
echter
van
meerdere
plantengeslachten
Deze
kevers
worden
oligofaag
genoemd
en
wanneer
zij
voedselplanten
van
meerdere
families
accepteren polyfaag.
Vaak
komt
het
voor
dat
bladkevers
als
volwassen
kever
meer
planten
eten,
terwijl
hun
larven
strikt
gebonden
zijn
aan
bepaalde
planten.
De
plantensoorten
die
alleen
door
volwassen
dieren
gegeten
worden,
worden
voedselplanten
genoemd;
de
planten
die
noodzakelijk
zijn
voor
hun
gehele
ontwikkeling
waardplanten.
Het
zich
beperken
tot
een
bepaalde
plant
heeft
voor-
en
nadelen.
Een
nadeel
is
dat
een
goudhaantje
altijd
de
plantensoort
zal
moeten
volgen.
Door
het
steeds
weer
aanpassen
van
plant
en
insect,
ontstaat
er
veelal
een
speciale
band.
De
plant
die
het
best
ontsnapt
aan
vraat,
zal
zich
het
meest
voortplanten,
en
de
plantensoort
past
zich
dan
aan.
Gespecialiseerde
herbivoren
moeten
volgen,
of
een
andere
plant
zoeken.
Gedurende
deze
evolutie
zijn
goudhaantjes
en
afweersysteem
van
de
planten
zo
verstrengeld
geraakt,
dat
de
speciale
stoffen
van
de
plant
noodzakelijk
zijn
geworden
in
hun
levenswijze.
Deze
genetische
verstrengeling
wordt
coëvolutie
genoemd,
waarbij
de
meest
complexe
relatie
tijdelijk
is
opgebouwd. (Zie ook Kuhn et al. (2004);
Termonia et al.
(2001)).
Ook in het voorkomen in het landschap, zie aldaar.
Als
een
herbivoor
eenmaal
een
plant
gevonden
heeft,
kan de plant niet
weglopen.
De plant kan zijn verdediging
alleen
ter
plekke
uitgevoerd
worden,
bijv.
met
haren,
met
het
ratelen
van
de
bladeren,
maar
vooral
met
chemische stoffen. Er zijn zelfs voorbeelden bekend waarbij de plant
"de hulp inroept" door de roofdieren van zijn herbivoren met een geur
te lokken.
 Niet
alleen
is
de
plantenafweerstof
op
zich
werkzaam,
maar
de
kever
vormt
de
afweerstoffen
om
tot
een
vluchtige
stof
en
gebruikt
deze
actief
voor
zijn
eigen afweer, zie overzichtsartikel (Strauss
et al., 2013).
Hiertoe
hebben
de
larven
een
dubbele
rij
klieren
op
hun
rug.
Bij
verstoring
stulpt
een
druppelachtig
blaasje
uit
een
klier,
waaruit
de
vluchtige
afweerstof
kan ontsnappen. De stof is vergelijkbaar met een wrattentinctuur,
bijvoorbeeld figuur hiernaast. De
larven
hebben
op
de
rugzijde
met
gepaarde
uitstulpbare
en
gespierde
blazen,
2
paar
op
borststuk
en
7
kleinere
op
achterlijf. Zie bijvoorbeeld foto's van Korzunovich, 2005 (Atlas
of the beetles of Russia) of de
FILM sigma1920
Japan 2009.
Bij Plagiosterna aenea verdedigen de larven zich door hun 'staart'
omhoog te doen (eigen waarneming 1991).
Niet
alleen
is
de
plantenafweerstof
op
zich
werkzaam,
maar
de
kever
vormt
de
afweerstoffen
om
tot
een
vluchtige
stof
en
gebruikt
deze
actief
voor
zijn
eigen afweer, zie overzichtsartikel (Strauss
et al., 2013).
Hiertoe
hebben
de
larven
een
dubbele
rij
klieren
op
hun
rug.
Bij
verstoring
stulpt
een
druppelachtig
blaasje
uit
een
klier,
waaruit
de
vluchtige
afweerstof
kan ontsnappen. De stof is vergelijkbaar met een wrattentinctuur,
bijvoorbeeld figuur hiernaast. De
larven
hebben
op
de
rugzijde
met
gepaarde
uitstulpbare
en
gespierde
blazen,
2
paar
op
borststuk
en
7
kleinere
op
achterlijf. Zie bijvoorbeeld foto's van Korzunovich, 2005 (Atlas
of the beetles of Russia) of de
FILM sigma1920
Japan 2009.
Bij Plagiosterna aenea verdedigen de larven zich door hun 'staart'
omhoog te doen (eigen waarneming 1991).
Goudhaantjes
gebruiken
de
speciale
brandstoffen
ook
als
herkenning
van
hun
waardplant. Read
(1969)
vermeldt dat
in
voorvoet
Chrysolina
soorten chemoreceptoren zitten. De
keuze
van
een
niet-giftige
plant
waarop
de
ouder(s)
hun
minder
mobiele
larven
achterlaten
is
van
groot
belang.
Zowel
de
voorkeur
om
eieren
te
leggen,
als
de
mogelijkheid
om
de
plant
te
kunnen
eten
zijn
genetisch
in
de
kever
vastgelegd.
Bacteriën
die
mogelijk
mee
kunnen
helpen,
worden
vaak
doorgegeven
doordat
de
ouder
hun faeces
over
de
eieren
verspreiden. Müller & Hilker
(2004) geven een overzicht van het onderzoek aan de ecologische betekenis
van faeces. Blum & Hilker
(2002) geven een algemeen overzicht van de manieren van chemische
verdediging van de eieren.
Nauwverwante
soorten,
die
op
twee
verschillende
planten
leven,
kunnen
hybride
vormen
die
beide
planten
eten
(Katakura
et al.
1989).
Een
verandering
in
voorkeur
voor
de
waardplant
zorgt
voor
scheiding
van
populaties,
doordat
de
kevers
elkaar
telkens
op
een
andere
plant ontmoeten. Gross (2001) onderzocht
populaties Chrysomela
lapponica uit Finland van Wilgen en populaties uit Tsjechië
van Berken. Op wilgen groeien de kevers het best, maar hebben zij last van
op wilg levende predatoren. De Zweefvlieg Parasyrphus wordt juist
aangetrokken door de afweerstof van wilgenkevers. Op berk groeien de
kevers minder goed, maar zijn in het voordeel doordat predatoren daar niet
komen. Kirst et al. (2011) tonen
aan dat de larven op berken de genexpressie voor afweerstof productie van
salicine hebben verloren. Het gevlekt wilgenhaantje Chrysomela vigintipunctata was
zeer zeldzaam in Nederland, maar recent vaker gevonden op Kraakwilg (Salix
fragilis L.) en Boswilg (Salix caprea L.). Everts (1922) meldt deze
bladkever van Bittere wilg (Salix purpurea L.) en Amandelwilg (S. triandra
L.). Als Chrysomela vigintipunctata zich met Schietwilg (Salix alba
L.) of Boswilg (S. caprea L.) hebben gevoed, hebben zij een hoge
sterftekans in de winter (Topp &
Beracz 1989). En zo worden er altijd soms onomkeerbare keuzes
(de evolutie) ingezet. Zie ook Termonia et al. (2001).
Aanpassen
Men
heeft
lang
gedacht
dat
de
fraai
gekleurde
dekschilden,
die
zo
kenmerkend
zijn
voor
deze
keverfamilie
uitsluitend
een
afschrikkende
functie
hebben
tegen
roofdieren.
Bladkevers
kunnen
de
afweerstoffen
van
planten
opslaan
en
daardoor
zelf
giftig
worden
voor roofdieren. Mohagheghi (2007)
geeft een uitgebreid overzicht van de synthese van afweerstoffen.
Opvallend
gekleurd
zijn
heeft
waarschijnlijk
ook
een
belangrijke
afschrikkende
functie,
maar
bij
de
Grote
hertshooigoudhaan
(Chrysolina
hyperici)
is
ontdekt
dat
de
goudgroene
dekschilden
als
een
kleurfilter
werken.
De
afweerstof
van
hertshooi
(hypericine)
is
fototoxisch,
dat
wil
zeggen
dat
het
werkzaam
wordt
in
het
zonlicht.
De
dekschilden
filteren
precies
de
kleur
uit,
die
hypericine
activeert.
Dit
werpt
een
heel
ander
licht
op
het
feit
dat
bij
veel
goudhaantjes
de
kleur
varieert
in
verschillende
landschappen,
en
dat
veel
larven
alleen
nachtactief
zijn.
In
het
algemeen
geldt
dat
blauwe
vorm
meer
voorkomt
in
bos
en
de
goudkleur
meer
in
het
open
veld.
Daarbij
moet
opgemerkt
worden
dat
goudhaantjes
in
de
loop
van
hun
levensjaren
steeds
metaalkleuriger worden en variatie in kleur kan komen door ander voedsel,
vocht, temperatuur of genetisch. Verschil in kleur kan vervolgens verschil
in fitness veroorzaken door diverse mechanismen, als seksuele selectie,
camouflage of waarschuwing tegen roofdieren, warmte absorptie e.d. (zie
ook Farrell & Sequeira 2004).
Dat
planten
ook
voordeel
hebben
van
gespecialiseerde
herbivoren,
is
vooral
bekend
bij
bestuiving.
Dan
is
er
een
directe
relatie
voor coëvolutie.
Bij
de
Chrysomelinae
is
dit
nauwelijks
bekent
soms
eten
kevers
pollen.
Of
het
verblijven
van
opvallend
felgekleurde
kevers
tussen
de
bloemen
enig
effect
heeft
op
andere
bestuivers
is
niet
onderzocht.
Doordat
een
aanpassing
in
eerste
instantie
lokaal
in
het
landschap
plaatsvindt,
komt
het
vaak
voor
dat
de
waardplant
geografisch
varieert.
Het
Groen
zuringhaantje
(Gastrophysa
viridula)
komt
tegenwoordig
in
het
laagland
optimaal
voor
op
ridderzuring,
maar
heeft
een
voorkeur
voor
alpenzuring
(Chevin
1968).
Rond
1900
zou
deze
kever
niet
in
Midden-Europa
zijn
voorgekomen,
maar
recent
(weer)
uit
het
gebergte
zijn
afgedaald
en
zijn
oude
voorkeur
voor Alpenzuring hebben
behouden.
In
Nederland
komt
het
Groen
zuringhaantje
ook
veel
voor
op
perzikkruid
en
is
bovendien
waarschijnlijk
nooit
weggeweest.
Aanpassen
aan
een
nieuwe
waardplant
gaat
bij
goudhaantjes
traag
(geschat
wordt
minimaal
14
generaties),
maar
deze
evolutie
is
binnen
een
mensenleven
waarneembaar. Natuurlijk beperkt een lichaam je gedrag, zie voor eetgedrag
Bieńkowski (2010),
en zijn de bladkevers sneller aan een andere plant gewend dan aan het type
weefsel dat gegeten wordt (Farrell &
Sequeira 2004). Het was (in 2003) bijvoorbeeld
te
verwachten
dat
de
Coloradokever
(ook
een
bladkever)
zich
gaat
aanpassen
aan
een
Europese
nachtschade.
De
aardappel
is
namelijk
niet
zijn
oorspronkelijke
waardplant.
In
1814
heeft
de
mens
de
aardappel
voor
het
eerst
in
het
leefgebied
van
de
kever
verbouwd
en
ongeveer
50
jaar
later
werd
de
kever
hierop
een
plaag.
In
Mexico
is
de
aardappel
is
in
1945
ingevoerd
en
de
Coloradokever
prefereert
daar
nog
steeds
de
inheemse
nachtschade.
Voor
de
Nederlandse
Coloradokevers
is
de
zwarte
nachtschade
uitermate
aantrekkelijk
om
eieren
op
af
te
zetten,
maar
de
larven
eten
deze
plant
niet.
Bitterzoet
zou
wel
een
goede
waardplant
kunnen worden (In 2011 werden larven massaal op Bitterzoet gevonden Calf
& Van Dam 2012).
Kevers
krijgen
van
Bitterzoet
hun
prikkel
tot
eileg
en
bij
de
larven
levert
de
giftige
cocktail
van
bitterzoet
in
de
darmen
geen
problemen
op.
Nu
is
het
wachten
op
de
set
genen
die
de
nog
onbepaalde
eigenschap
vormen,
waardoor
zij
er
nog
niet
op
zitten.
Bijvoorbeeld
de
biologische
klok.
Dit
zou
kunnen
door
het
biologisch
ritme
van
de
plant
"te
lezen".
Planten
produceren
niet
in
elk
seizoen
dezelfde
stoffen.
Een
insect
kan
hierdoor
zelfs
het
seizoen
proeven!
Coloradokevers
die
gevoed
worden
met
oud
loof
stoppen
met
eileg
en
gaan
in
winterslaap
(De
Wilde
& Ferket
1967). Voor een ander groot deel is het aanpassen aan het ontsnappen
aan predatoren, een dilemma met voedsel. Chrysomela
lapponica was ook even in Nederland vrij
algemeen, door een verandering Wilg (Salix) naar Berk (Betula),
weg van de predatoren, en kregen door dit voedsel te maken met een hogere kans
op sterven in de
winter (Topp & Beracz 1989). Bij
de larven hebben Kirst et al. (2011)
aangetoond dat op Berk (Betula) het gen voor afweerstof productie met salicine
niet meer tot expressie komt, wat natuurlijk voordeel is bij adequate
milieu-eisen.
De
kans
op
de
mogelijkheid
van
vermenging
is
meestal
afhankelijk
van
de
tijdsduur
van
scheiding, dat is een manier waardoor soorten
ontstaan.
Voor
de
montane
Oreina
cacaliae
is
vastgesteld
dat
populaties,
die
3
km
van
elkaar
gescheiden
zijn,
genetisch
verschillen.
En
volgens
berekening
zou
op
200
km
afstand
geen
genetische
uitwisseling (gene-flow)
meer
mogelijk
zijn
(Knoll
et al.,
in
Jolivet
& Cox
1996). Maar bij Chrysomela
tremula stellen Génissel
et al. (2000): "
The major
result of this study is the finding that there are no obvious limitations to
gene flow between populations of C. tremulue over large geographical distances (several
hundreds of kilometres)". Daarentegen bij
Plagiodera versicolora
is
vastgesteld
dat
zij
op zeer korte
afstand
genetisch kunnen verschillen,
zelfs
tussen
twee
solitaire
bomen
(McCauley
et al.
1988).
De
eerste
is
een
slechte
verspreider
(ongeveer
50
m
per
jaar)
en
de laatste twee vliegen
goed.
Zelfs
tussen
aparte
soorten
is
vermenging
niet
uit
te
sluiten.
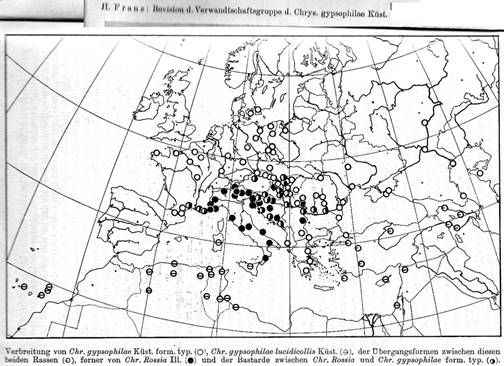
Van
Chrysolina
subgenus
Stichoptera
zijn
door
Franz
(1938)
diverse
hybride
gebieden
beschreven.
Deze
stelling
is
weinig
onderbouwd,
maar
het
is
buiten
kijf
dat
er interspecieke
gene-flow bestaat.
Van
deze
roodgezoomde
Goudhanen
is
het
onderscheid
echter
zeer moeilijk. Recent is er zelfs een
nieuwe soort beschreven van de Atlantische kust van Frankrijk, zie Bourdonné; Doguet & Petitpierre (2013).
C.
gypsophilae
wordt
bijvoorbeeld
gemeld
van
de
Canarische-Eilanden
tot
Scandinavië
met
een
type
locatie
in
Italië.
Doordat
er
een
band
in
de
verspreiding
lijkt
te
zijn
is
dit
mogelijk
een
hybride
tussen
C.
kuesteri
en
C.
sanguinolenta,
vergelijk
andere
taxa
(Hewitt
1999).
Voor
Nederland
lijkt
onderzoek
aan
deze
complexiteit
niet
meer
zinvol,
daar
zij
alle
op
de
meeste
plaatsen
zijn
verdwenen.
Dit
is
wel
het
geval
bij
algemene
of
schadelijke
soorten.
Aan
aanpassingen
die
overgedragen
worden
door
dispersie
is
nauwelijks
onderzoek
gedaan.
Daarbij
moet
men
denken
aan
het
aantal
generaties
per
jaar;
allerlei
ritmen
in
activiteit,
aanpassingen
aan
menselijke
bestrijding,
predatie, enz.;
voedsel-
en
waardplanten
enz.
[KRAF95].
Gomez-Zurita,
Funk & Vogler (2006) stellen de hypothese op dat Calligrapha in
Noord-Amerika door parthenogenese zich heeft kunnen verspreiden vanuit
Midden-Amerika . Deze aseksuele voortplanting zou veroorzaakt zijn door
interspecieke hybridisatie. In Midden-Amerika leven de soorten van dit
geslacht op kruiden en de 38 soorten in Noord-Amerika op struiken en
bomen. De soorten komen vaak massaal voor langs rivieren op o.a. Els (Alnus),
Wilg (Salix), Kornoelje (Cornus) en Iep (Ulmus).
Vooral de soortenrijkdom in Oost-Canada (Ontario en Quebec) is groot. Van
parthenogenese is bekend dat het een groot verspreidingsvermogen kan
opleveren, maar kan natuurlijk leiden tot een evolutionair doodpunt.
Chrysolina
polita,
die
het
Moertje
genoemd
wordt,
is
de
algemeenste
van
het
geslacht.
Het
succes
van
deze
soort
blijkt
bij
de
vliegensvlugge
kolonisatie.
De
kever
is
oligofaag
binnen
de
lipbloemenfamilie
en
komt
optimaal
voor
in
moeras,
bijv.
op
wolfspoot.
Buiten
het
moeras,
in
suboptimaal
gebied,
op
dijken
en
in
bosranden
eet
het
Moertje
een
andere
lipbloem:
hondsdraf.
Weer
in
een
ander
moeras
is
een
grote
populatie
op
munt
te
vinden.
Hondsdraf
is
weliswaar
een
waardplant
van
het
Moertje,
maar
slechts
tussen
twee
optimale
biotopen.
Dit
houdt
voor
het
Moertje
de
mogelijkheid
van
reizen
in
en
daarom
noem
ik
het
in
't
veld
een reisplant.
Het
is
niet
definitief,
het
is
slechts
op
dit
moment
de
rol
van
de
hondsdraf
in
de
huidige
levenswijze
van
het
Moertje,
maar
het
belang
van
reisplanten
is
in
elk
landschap
te
zien.
Van
meerdere
soorten
zijn
dergelijke
reisplanten
te
onderscheiden:
voor
de
Grote
goudhaan
(C.
graminis)
is
dit
boerenwormkruid
langs
rivieren
totdat
de
kever
grote
lipbloemigen
in
een
stabiel
moerasbos
tegenkomt.
Biogeografie
De
waardplanten
van wilgenhaantjes
komen
vooral
voor
in
het
stroomdal
van
rivieren
en
staan
vaak
periodiek
in
het
water. Wilgenhaantjes
kunnen
ten
opzichte
van
andere
goudhaantjes
goed
vliegen.
Het
voorkomen
van
de
verschillende
soorten
wordt
vooral
bepaald
door
het
stadium
waarin
een
droog
habitat
nodig
is.
Griendhaantjes
(Phratora-sp.)
overwinteren
in
holtes
van
de
stam,
zodat
's
winters
en
in
het
voorjaar
de
bomen
onder
water
kunnen
staan.
Zij
volbrengen
echter
hun
popstadium
in
de
bodem
onder
de
boom.
In
de
zomer
moeten
de
bomen
waarop
zij
leven
dus
droog
komen
te
staan.
Het
Rond griendhaantje of Veelkleurig
wilgenhaantje (Plagiodera versicolora)
en
Chrysomela-soorten
kleven
hun
pop
aan
het
blad.
Zij
kunnen
bomen
op
hun
menu
zetten,
die
zomers
ook
onder
water
staan. Door het onder water staan van de bomen kunnen de larven ook
voedselgebrek krijgen en gaan dan synchroon verpoppen (LeSage
1996), met een massavlucht tot gevolg. 's
Winters
moeten
de
Chrysomela's
echter
naar
een
hoger
gelegen
landschap
vliegen,
want
overwinteren
doen
zij
onder
bladeren
in
droge
bosbodem.
Het
kleine
monofage
Rond griendhaantje heeft
drie
tot
vier
generaties
per
jaar
en
kan
zijn
aanpassingen
waarschijnlijk
daardoor
snel
doorgeven.
Voor
hun
overwintering
zijn
zij
veel
minder
kieskeurig
en
vallen
gewoon
in
de
herfst
met
de
bladeren
mee.
De
roodbruine wilgenhanen
van
het
geslacht
Gonioctena
moeten
voor
zowel
hun
pop
als
voor
hun
overwintering
naar
droge
bodem.
Zij
leven
meer
in
droge
bossen
en
verspreiden
zich
over
het
algemeen
langzaam.
In de duinen en het
laagveen ontbreken zij, terwijl hun
waardplanten daar wel voorkomen. Een
plant als voedsel is niet genoeg, de larven moeten het ook kunnen eten: de
waardplant, maar het voorkomen van de waardplant verschilt met die van de
bladkever en dus komt daarbij nog het landschap.
Goudhaan-soorten
(Chrysolina-sp.)
leven
op
kruiden
en
vliegen
praktisch
nooit,
zij
lopen
vooral
langs
lijnvormige
landschapselementen.
De
meeste
Nederlandse
dieren
hebben
weliswaar
volledige
vleugels,
maar
hun
vliegspieren
zijn
gereduceerd.
Met
zekerheid
zijn
alleen
Chrysolina
graminis en C.
americana
vliegend
waargenomen. Door de waarnemingen van J.-C. Bourdonné is voor de Franse
situatie het aantal inmiddels flink gegroeid (Winkelman
& Debreuil 2008 en Winkelman 2013).
De
Coloradokever
heeft
zich
20e
eeuw
met
gemiddeld
60
km
per
jaar
naar
het
noorden
uitgebreid.
Deze
grote
exotische
soort
breidt
zich
vooral
vliegend
uit.
Van
onze
inheemse
goudhaantjes
vliegen
de
soorten
van
het
geslacht
Chrysomela
het
beste.
Bij
de
recente
invasiegolf
van
Chrysomela
lapponica,
die
Beenen
(1985)
beschrijft,
komt
de
uitbreiding
ongeveer
uit
op
10
km
per
jaar.
In
1990
vond S.C.
Langeveld
hem
zelfs
al
in
de
Amsterdamse Waterleidingduinen.
De
lopende
goudhanen
(Chrysolina-sp.)
komen
niet
verder
dan
100
meter
per
jaar.
Samen
met
de
vleugelloze
Reuzenhanen
(Timarchinae)
zijn
goudhanen
de
slechtste
verspreiders,
die
vooral
de
zoomen
van
landschappen
bewonen.
Het
betreft
hier
ware
olifanten
op
schaal
1:500,
met
rudimenten
van
vleugels.
En
voor
deze
lopende
dieren
gaat
passieve
verspreiding,
meegevoerd
worden
met
rivierwater,
vele
malen
sneller. In het hooggebergte kennen de soorten andere problemen, daar
moeten de soorten steeds hogere delen bewonen door de huidige
klimaatsverandering totdat zij uitsterven, zie het voorbeeld uit het
oosten van het Californische Sierra Nevada gebergte, waar Chrysomela
aeneicollis voorkomt op 2700-3600 m hoogte (Dahlhoff
et al. 2008).
De Roodgezoomde
duingoudhaan
(Chrysolina
limbata), loopt ook met z'n DNA te zeulen en leeft
in
droge
graslanden
op een Weegbree (Plantago),
maar
toch
is
zijn
huidige
beperkte
voorkomen
in
de
Hollandse
duinen door
de
loop
van
de
rivieren
bepaald
(Winkelman
1993).
Op
graslanden
van
de
voormalige
rivierduinen
leefde
deze
soort
waarschijnlijk
ook.
De
Reuzenhanen
bewonen
meer
de
heuvels,
maar
maken
bij
hun
uitbreiden
ook
gebruik
van
de
rivier,
wat
aan
hun
voorkomen
is
te zien.
Behoudens
één
vondst
in
Den
Haag,
komen
de
Reuzenhanen
tegenwoordig
stroomafwaarts
niet
meer
voor
(Beenen
1988).
Of
de
vleugelloze
Timarcha
ooit
zelf
het
duinbos
heeft
bereikt
is,
met
zo'n
aaibare
soort,
niet
zeker.
De
recente
waarneming
uit
Den
Haag,
staat
echter
niet
op
zich:
de
soort
is
vroeger
ook
al
in
Leiden
en
Den
Haag
gevonden.
In
de
laatste
IJstijd
is
Noordwest-Europa
bijna
geheel
bedekt
geweest
met
ijs.
Europa
kende
beperkte
uitwijkmogelijkheden
in
de
laatste
IJstijd.
Gematigde
soorten werden
tot
ver
achter
de
ijsgrens
gedreven,
naar
het
land
langs
de Middellandse-Zee.
Echte
warmteminnende
soorten
naar Noord-Afrika.
Veel
soorten
waren
door
de
koude/droogte
geïsoleerd geraakt.
Door deze gebeurtenissen,
als
IJstijden,
destructie
van
landschappen
door
de
mens,
overstroming,
verdwaling e.d.,
raken
groepen
kevers
geïsoleerd
van
hun
verwanten.
Hun
verwanten
kunnen
zich
anders
ontwikkelen.
Soms
veranderen
zij
zodanig
dat
zij
niet
meer
met
elkaar
kunnen
reproduceren.
Sommige
soorten
veranderen
sneller
dan
anderen.
Timarcha
ken
al
separatie
op
het
Kanaal-Eiland
Chaussy
dat
pas
sinds
1200
een
eiland
is
(Bechyně
1950). Ook in het grote Australië zijn bijna onvoorstelbare kleine
isolatie processen bekend, zoals een soort van Canobolas Reid,
Jurado-Rivera & Beatson 2009 die natuurlijk beschreven is van 1
exemplaar. Van deze type locatie is volgens Reid
et al. (2009) slechts
bekend: "could be applied to a broad area of limestone gorge country
from Jenolan Caves (800m) to Jenolan State Forest (1200m)".
Laatglaciaal
luiden
vooral
de
pioniers
onder
de
Composieten
het
einde
van
de
IJstijd
in
(Van
Huissteden
& Ran
1993).
Ongeveer
13.000
jaar
geleden
is
er
een
toename
van
Alsem
(Artemisia).
Alsem
is
de
waardplant
van
Chrysolina
(Chalcoidea)
en
mogelijk
ook
van
Chrysolina
(Craspeda),
deze
zijn
alle
donker
van
kleur
en
roodgerand.
Overblijfselen
van
deze
levensgemeenschap
zijn
zelden,
en
nooit oorspronkelijk. De Berk
en de
Ratelpopulier waren de
eerste
bomen. Voor meer
soorten
van
verschillende
genera
geldt
dat
de
soorten
die
in
het
Arctisch
gebied
voorkomen,
als
laatglaciaal
fossiel
bekend
zijn
(Lemdahl
& Gustavson
1997).
Zij
zijn
vaak
scherp
en
opvallend
getekend,
bijvoorbeeld
Hydrothassa
hannoveriana
en
P.
phellandrii. Van
Chrysolina (Anopachys)
is de soort relucens (Rossenhauer, 1847) een
voorbeeld van een noord-zuid disjuncte: in de Alpen (Italië en Oostenrijk)
en Noord-Azië (Kippenberg 2010)
en Karelië Karjala Karelija Карелия
(Silfverberg 2004).
In
veel
genera
vicariëeren
soorten
in
het
Palearctische
en
Nearctische region,
en
slechts
enkele
worden
als
één
soort
beschouwt
met
een
holarctische verspreiding, zie ook (Elias
& Crockera 2008).
Hun
huidige
status
is
taxonomisch
weergegeven
naar
de
laatste
inzichten.
Gastrophysa
polygoni
en
meerdere
Phaedon-soorten
zijn
echte
holarctische
soorten.
Volgens
(Lindroth
1957)
is
Chrysolina
staphylaea trans-Atlantisch verspreidt,
over
de
landbrug
van
Europa
(Schotland)
via
IJsland
naar
Amerika.
Deze landbrug
zou
tot
in
het
Tertiaire
Plioceen
en
het
Quartaire
Pleistoceen
hebben
voortgeduurd.
Daarnaast
zijn
er
vicariëerende
soorten,
soorten
die
opgesplitst
zijn
door
geografische
scheiding:
Chrysomela
saliceti
is
verwant
met
de
Nearctische
C.
crotchi
Brown,
1956; Chrysomela
heeft
een
duidelijkere
vicariantie
in
het
Arctische
gebied,
dan
Gonioctena.
Phratora
polaris
met
P.
hudsonia
Brown,
1951
en
P.
frosti
Brown,
1951.
En verder de recent door de mens geïmporteerde soorten
als de schadelijke Plagiodera
versicolora
en biologische bestrijders uit Chrysolina.
In
Nieuw
Zeeland
en
Australië
(PAC,
AUS
en NZL) meer dan 40
genera
met
veel
endemische
Chrysomelinae
(vooral
tribus
Paropsini). Voor de
Australische fauna is er een recente revisie van Reid
(2006) voor de 750 soorten. Zij
komen
vooral
voor
op
jonge
planten
van
de
tropische
Myrtaceae
en
Vlinderbloemen (Fabaceae).
De
kenmerkende
Paropsis
Olivier
1807
op
jonge
Eucalyptus
L'Héritier
de
Brutelle (Myrtaceae)
en
Calomela
Hope,
1840
op
Acacia
Miller (Fabaceae) (Selman
1979) (zie  literatuur). Zij
vertonen
waarschijnlijk
een
functionele monofagie,
want
hun
verwanten
in
het
Oriëntaalse
region
eten
Rosaceae (Jolivet
& Hawkeswood 1995).
literatuur). Zij
vertonen
waarschijnlijk
een
functionele monofagie,
want
hun
verwanten
in
het
Oriëntaalse
region
eten
Rosaceae (Jolivet
& Hawkeswood 1995).
Ook
komen
ze
voor
in
Nieuw-Zeeland
dat
veel
op
de
Australische
fauna
lijkt. Jurado-Rivera et al. 2008
onderzochten de evolutionaire relatie met hun waardplanten.
In
het
Pacifische
region
komen
zij
niet
voor
op
de
oceanische
eilanden,
maar
uitsluitend
op
continentale
eilanden:
de
Fidzji-Eilanden
en
West
Samoa
(Gressitt
1957). Op Nieuw Caledonië één endemisch genus met één bekende
soort, Zira nitens Reid & Smith, 2004 (Jolivet
& Verma 2009). Chalcolampra octodecimpunctata
(Fabricius) en Chrysophtharta cloelia Stål (= Phola
Wse) zijn geïmporteerd, de eerste voedt zich met Eucalyptus
sp., de tweede met Vitex
sp. (Jolivet & Verma 2002).
In
het
Antarctische
region
2
genera
op
Tierra
del Fuego,
de
Magellan
Archipel
en Patagonië.
Deze
vertonen
overeenkomsten
met
Australische
en
Afrikaanse
genera.
Een
hiermee
verwant
genus
in
Australië
leeft
op
de
Antarctische
beuk
(Nothofagus).
Deze
bomen
zijn
een
Tertiairrelict,
zij
komen
zowel
in
Australië
als
in
Zuid-Amerika
voor.
Het
voorkomen
van
verwante
Chrysomelinae
in
Australië,
Zuid-Amerika
en
Afrika
duidt
op
een
ontstaan
van
de
onderfamilie
in
de
Midden-Jura
toen
Gondwanaland
nog
bestond.
Oriëntaalse
fauna (ORI)
is
weinig
vertegenwoordigd
met
4
genera
en
zonder
endemische
genera.
Vooral
Phyllocharis
Dalman,
1824
op
de
planten
uit
de
IJzerhardfamilie (Verbenaceae) en
Chalcolampra
Blanchard,
1853
op
Helmkruiden (Scrophulariaceae)
en
Lipbloemigen (Lamiaceae). Zij dringen zelfs door in het Australische region (Reid
2006; Mohamedsaid 2009).
In Noord-Amerika (NEA)
geen
endemische
genera.
In
totaal
15
genera,
130
soorten,
waarvan
de
oorspronkelijk
Neotropische
Calligrapha
Chevrolat
in Dejean,
1836
het
grootste
genus
is
met 38 soorten (zie ook Gómez-Zurita
et al. 2004).
Zij
komen
nog
in
Zuid-Amerika
voor,
vliegen
goed
en
eten
specifieke
struiken
en
bomen
van allerlei families, zie ook hierboven.
Zuid-Amerika (NEO)
heeft
het
grootst
aantal
soorten
in
31
genera,
waarvan
Doryphora
Illiger,
1807
het
opvallendste
genus is (zie ook
Eberhard 1981 en Windsor
et al. 2013)
In
Afrika (AFR)
zijn
er
slechts
19
genera,
waarvan
vele
endemisch.
In
tegenstelling
tot
Zuid-Amerika
zijn
zij
uit
Afrika
nauwelijks bekend
uit
het
oerwoud.
De
bekendste
zijn
Chrysolina
L.
en
Ceralces
Gerstaecker,
1855.
De
genera
van
Madagaskar
zijn
verwant
met
de
genera
van
het
vaste
land.
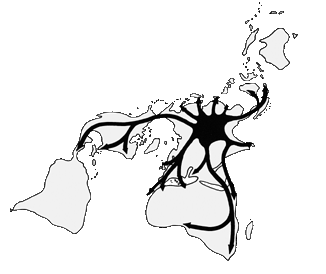
Dispersie van het genus
Chrysolina
volgens
Daccordi (1996)
geprojecteerd
op de huidige wereldkaart.
Bedreiging
Van
de
Bladkevers
zijn
soorten
van
het
geslacht
Timarcha
unieke
levende
fossielen.
Zij
lopen
erg
traag,
kunnen
niet
ontsnappen
door
op
te
vliegen.
De
dieren
hebben
rudimentaire
vleugels
en
hun
pekzwarte
dekschilden
zijn
als
onze
fontanel
vergroeid.
De
larven
van
onze
Reuzenhanen
hebben
kennelijk
veel
warmte
nodig
en
zijn
ook
zwart
en
zijn
uitsluitend
midden
op
de
dag
in
de
zomer
actief.
In
Noord-Amerika
zijn
zij
echter
nachtactief.
Hun
majestueuze
verschijning
kan
inderdaad
vergeleken
worden
met
de
Atheense
rechters:
de Timarches.
Hun
voorkomen
wordt
bedreigd
door
verlies
en
versnippering
van
hun
woongebied.
Langs
de
Middellandse
Zee
heeft
de
vakantiecultuur
zijn
tol
geëist.
Endemische
soorten
van
eilanden
in
het
Mediterrane
subregion
zijn
reeds
uitgestorven.
Het
inkrimpen
van
het
areaal
van
Timarcha
binnen
Europa
lijkt
in
de
onze
tijd
Nederland
te
passeren.
Verspreiden
kunnen
zij
niet
meer
en
geïsoleerde
populaties
sterven
langzaam
uit.
Vroeger
mogelijk
het
hele
Pleistocene
district,
het
rijnduin
district
daarbij
inbegrepen,
het
langst
gehandhaafd
in
het
Fluviatiel
district
en
tegenwoordig
alleen
in
het subcentrope-
en
het
Krijtdistrict
(Beenen
1990).
Meer soorten en het gehele geslacht Gonioctena,
ontbreken op de Nederlandse duinen. Maar voor enkele populaties zijn
de
duinen juist een
relict
situatie.
Voor
meer
soorten
bevindt
zich
tegenwoordig
juist
in
de
duinen
de
grootste populatie (of de enige, bijvoorbeeld Chrysolina
limbata). Enerzijds
zijn
de
duinpopulaties
geïsoleerd
en
hebben
een
grote
kans
uit
te
sterven,
maar
anderzijds
kan
een
soort
in
isolement
het
rijk
alleen
hebben
doordat
concurrenten,
specialistische predatoren,
ziekten e.d.,
ook
moeite
hebben
de
duinen
te
vinden.
Zo
komt
het
Gezoomd wilgenhaantje
(Chrysomela
collaris)
tegenwoordig
bijna
uitsluitend
voor
in
de
duinen.
Het
gezoomd wilgenhaantje
is
monofaag
en
waarschijnlijk
erg
trouw
aan
zijn
biotoop:
kruipwilg
in
plassen.
Daartegenover
komt
de
oligofage
Kleine
populierenhaan
(C.
saliceti)
tegenwoordig
nog
wel
in
het
binnenland
voor
maar
(nog)
veel
minder
op
de waddeneilanden.
Chrysomela's
zijn
goede
vliegers,
die
zelfs
over
het
veenweide
gebied
de
duinen
en
de
waddeneilanden
kunnen
bereiken.
Dit
gebeurt
waarschijnlijk
periodiek,
met
golven,
als
de
populatie
tijdelijk
enorm
groeit,
zoals
we
dit
meemaken
met
C.
lapponica
(Beenen
1985) en prompt daarna vanaf 1985 en wederom vanaf 1993 met Chrysomela
vigintipunctata. Voor alle soorten is per provincie de presentie aangeven,
vóór en na 1966, dus vóór en na Brakman (1966), zie daarvoor het overzicht
of klik op de Nederlandse kaart (op de kaart wordt echter vóór en na
1950 gehanteerd).
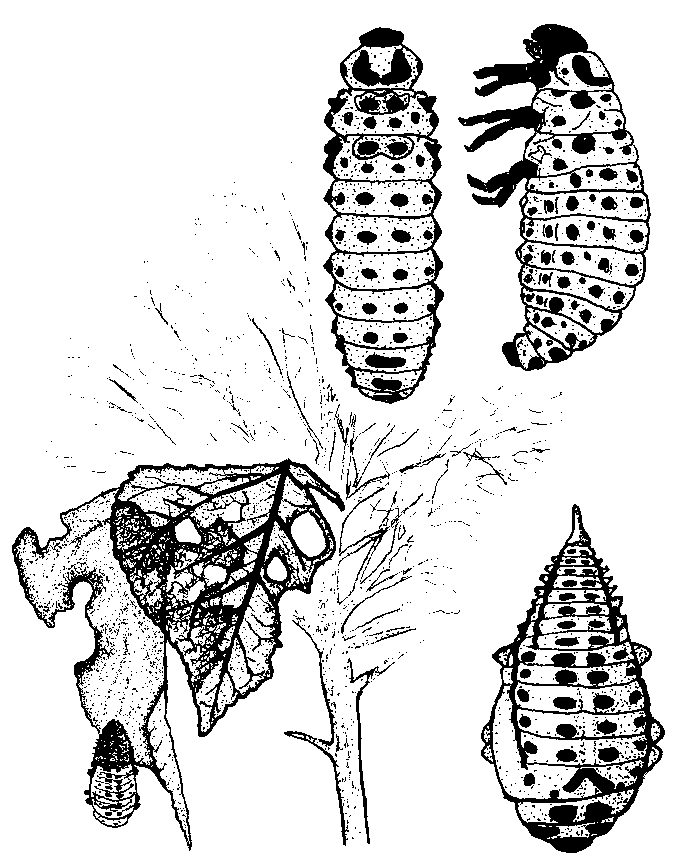



Chrysomela soorten.
Wat
zou
men
hiervan
kunnen
zien
in
't
veld?
Ten
eerste
kijkt
men
wat
de
kever
doet:
eten,
rusten,
paren,
op
de
larven
passen
enz.
En
hoe
is
het
lokale micro-milieu:
op
welk
uur
van
de
dag
is
er
zon,
en
hoeveel
water
is
er
aanwezig
en
wat
is
de
aard
van
de
bodem?
Ten
slotte
waar
bevinden
zich
de
larven,
eieren
en
de
overige
kevers,
en
wat
doen
zij? Het bepalen van de stukjes landschap met hun waardplant(en), en welke
noodzakelijk zijn voor hun leven en het inzien dat deze levensgemeenschap
allemaal tijdelijk kan zijn, vormt een hele studie.
Zelfs
bijvoorbeeld in Noord-Hollandse Enkhuizen is er iets te zien (22-7-2011 waarneming.nl-62004025).
In
het
voorjaar
vond
ik
in
een
dras/nat
graslandje
langs
het
Gein
bij
Abcoude
larven
van
het
Gezoomd
moerashaantje
(Hydrothassa
marginella)
etend
aan
onbehaarde
boterbloemen.
Na
een
speurtocht
naar
volwassen
dieren
vond
ik
er
een
tiental
in
de
droge
wegberm
etend
aan
een
behaarde
boterbloem.
Dergelijke
waarnemingen
leiden
tot
vragen
en
ideeën:
zijn
de
gevonden
volwassen
kevers
de
ouders
van
de
larven?
Gaan
volwassen
kevers
na
eileg
reizen,
en
ontmoeten
zij
elkaar
aan
de
rand
van
hun
biotoop?
Uit
de
literatuur
is
bekend
dat
larven
hun
eierenleggende
ouders
met
chemische
stoffen
verjagen.
Kweken
Er
zijn
soorten
die
zo
verborgen
leven
dat
nodig
is
om
ze
te
leren
kennen.
Hieronder
zal
ik
kort
mijn
waarnemingen
aan
de
Paarse
goudhaan
(Chrysolina
sturmi)
beschrijven.
Op
een
zonnige
dag
in
de
zomer
vond
ik
de
Paarse
goudhaan
in
het
binnenduinbos.
Zij
was
hoog
in
een
bijvoet
geklommen,
welke
in
de
halfbeschaduwde
berm
groeit
naast
een
schietterrein.
Ik
besloot
het
actieve
dier
mee
te
nemen.
Een
bakje
zand
met
een
plantje
geplaatst
in
een
kweekbak
vormde
zijn
versimpeld
biotoop.
De
eerste
week
was
de
kever
erg
actief
in
het
klimmen
en
lopen.
Welk
voedsel
zou
deze
goudhaan
nodig
hebben?
De
meeste
goudhanen
eten
lipbloemigen,
maar
het
stukje
bosandoorn,
dat
ik
van
de
vindplaats
meegenomen
had,
werd
niet
gegeten.
Na
het
aanbieden
van
hondsdraf
uit
Amsterdam,
groef
de
kever
zich
net
zichtbaar
in
het
zand.
De
kever
was
overdag
meestal
verscholen
en
kwam
in
de
loop
van
de
lange
zomeravonden
te
voorschijn.
Bij
zonsopkomst
was
de
kever
bijna
altijd
weer
teruggekeerd.
Het
gebeurde
ook
dat
de
kever
zich
dieper
ingroef
en
zich
enkele
dagen
niet
liet
zien.
In
september
vermoedde
ik
dat
de
kever
in
winterslaap
was
gegaan,
maar
toen
ik
haar
opzocht
in
de
surrogaat
bodem,
zag
ik
daar
15
larfjes
rond
kruipen.
Buiten
had
het
flink
geregend
en
de
hondsdraf
had
zich
uitgebreid.
Maar
wanneer
was
de
kever
bevrucht?
Het
is
bekend
dat
sperma
opgeslagen
kan
worden,
maar
ook
dat
vrouwelijke
kevers
tot
aseksuele
voortplanting
overgaan
als
er
geen
mannetje
komt.
De
larven
vertoonden
een
grote
individuele
variatie
in
activiteit,
maar
over
het
algemeen
aten
zij
ook
's
nachts.
Zij
klommen
langs
de
stengel
omhoog
en
lieten
in
de
ochtend
een
karakteristiek
vraatspoor
achter.
Tijdens
de
koude
winternachten
waren
de
larven
bij
4
°
C
actief.
Er
was
er
een
grote
variatie
in
de
ouderdom
van
de
larven
tijdens
de
winter,
want
tot
in
januari
heeft
de
kever
eieren
gelegd.
Bladkevers
zijn
een
uitstekende
diergroep
om
het
landschap
biologisch
te
bestuderen,
door
hun
variatie
in
biotoop,
hun
variatie
in
talrijkheid,
maar
vooral
doordat
de
wisselwerking
met
planten
in
het
veld
zo
zichtbaar is (zie ook Staines
& Staines 1998).
zeldzaamheid
en
bedreiging
Op
grond
van
het
aantal
exemplaren
in
de
collectie
van
het
Zoölogisch
Museum
te
Amsterdam,
het
aantal
provincies
welke
genoemd
zijn
in
Brakman
(1966)
en
de
gegevens
uit
de
Nederlandse
literatuur,
zijn
de
soorten
ingedeeld
in
6
klassen
van
talrijkheid:
zeer
zeldzaam,
zeldzaam,
vrij
zeldzaam,
niet
algemeen,
algemeen
en
zeer
algemeen.
Voor
de
soorten
welke
in
de
eerste
4
klassen
vallen,
is
deze
klasse
ook
omgezet
in
een zeld(zaamheids)code.
Zeer
zeldzaam:
code
0,
zeldzaam:
code
1,
vrij
zeldzaam:
code
2,
niet
algemeen:
code
3.
Van
de
zeer
zeldzame
soorten
en
van
soorten
waarvan
de
verspreiding
in
Nederland
bekend
is,
is
de
mate
van
bedreiging
vastgesteld.
In
april
1994
is
een
oproep
om
gegevens
te
verkrijgen
geplaatst
in
het
mededelingenblad
van
de
sektie
Everts
van
de
Nederlandse
Entomologische Vereniging (www.nev.nl
PDF).
De
Rode
Lijst
is
opgesteld
uitgaande
van
zijn
in
voorkomen
in
de
twee
helften
van
de
20e
eeuw,
geclassificeerd
volgens
tabel
1
en
naar
de
IUCN
categorieën
benoemd
op
de
volgende -ongetwijfeld verouderde- manier (zie ook IUCN 2001):
categorie
0: RE uit
Nederland
verdwenen;
na
1950
niet
meer
waargenomen;
categorie
1: CR op
het
punt
van
verdwijnen;
de
soort
is
na
2000
waarschijnlijk
uit
Nederland
verdwenen;
categorie
2: EN sterk
bedreigd;
de
soort
is
na
2050
waarschijnlijk
uit
Nederland
verdwenen;
categorie
3: VU bedreigd;
de
overige
zeer
zeldzame
en
afnemende
soorten.
| |
25 - 50 % |
51
- 75 % |
> 75 % |
| 1 - 2 |
2 |
1 |
1 |
| 3 - 7 |
3 |
2 |
1 |
| 8 - 20 |
3 |
3 |
2 |
TABEL
1:
Indeling
in
categorieën
van
bedreiging.
Aantal recente UTM-GRID's,
versus
het
percentage
afname.
 29
vii 2010: De
overgebleven Timarcha's komen zo niet
meer op de Rode Lijst, zie ook waarnemingen.be
; waarneming.nl
en waarnemingen.be
wellicht is de grens van 20 UTM te licht.
29
vii 2010: De
overgebleven Timarcha's komen zo niet
meer op de Rode Lijst, zie ook waarnemingen.be
; waarneming.nl
en waarnemingen.be
wellicht is de grens van 20 UTM te licht.
21 xii 2010: Van de hele familie
zijn een veertigtal van de 315 Nederlandse soorten blad- en zaadkevers ruim 50
jaar niet meer gevonden, ongeveer 13 % (Beenen
& Winkelman 2010).
1
XII 2015: Vergeleken met de extinctie van het ouder leven in het
Antropoceen (als koralen Edward O. Wilson, 2016 Half-Earth),
komt de gehele familie, als jonge telg in de evolutie er goed van af (Smith
& Marcot 2015)
Alleen
categorie
4
(potentieel
bedreigd) NT is
vervangen
door
de
zeldzaamheidscode
0.
Bij
het
vergelijken
van
het
voorkomen
in
de
2
perioden,
is
als
maat
de
presentie
in
een
UTM-GRID
gebruikt.
Hiermee
is
een
eventueel
verschil
in
verzamelintensiteit
van
deze
perioden
gecorrigeerd,
wat
de
uitspraak
over
de
verandering
in
voorkomen
onnauwkeuriger
maar
wel
betrouwbaarder
maakt.
Gonioctena
pallida
is
ook
zeer
zeldzaam,
maar
om
de
verspreiding
van
deze
soort
te
onderzoeken
zouden
van
alle
soorten
van
Gonioctena
subgenus
Goniomena
het
genitaal
bekeken
moeten
worden
en
hiervoor
ontbrak
het
mij
aan tijd (DD).
Chrysolina
americana
was
in
Nederland voor 1992 ook
zeer zeldzaam (2 records), maar
deze
soort leeft
in
antropogeen
milieu
en
is veel algemener (2012: 115 records). Mogelijk zijn
de
soorten
met
een
zeldzaamheidscode
ook
bedreigd,
bijvoorbeeld
Chrysolina
fuliginosa welke mogelijk in het jaar 2078 verdwenen is; Hydrothassa hannoveriana
(zie Williams 2015);
Phaedon
concinnus en
Chrysomela
tremula
(na 1966 nog maar in 1 provincie, daarvoor in 7 provincies).
Van
Chrysolina
gypsophilae
is slechts één vondst bekend van 1958. Zie daarvoor het kaartenoverzicht
en ook het handige
overzicht voor alle 4163 kevers met het boek : Catalogus van de
Nederlandse kevers (Vorst 2010) of
voor deze groep 59 kevers op deze site in het provincie
overzicht.
Er
is
uitgegaan
dat
een
soort
ook
aanwezig
was
in
de
voorafgaande
periodes,
omdat
bij
zeldzame
soorten
de
kans
groot
is
dat
er
onderbemonsterd
is.
TABEL
2:
overzicht
van
de
zeldzame soorten, en de algemene Chrysolina fastuosa als blanco.
AFKORTINGEN: vnd:
totaal
aantal
vondsten
(inclusief
unieke
opgave
in
de
literatuur); ex.:
totaal
aantal
gevonden
exemplaren, unieke (literatuur)
opgave
zijn
op
1
exemplaar gesteld;
laatste:
jaar
waarin
de
laatste
vondst
gedaan
is;
At-2:
aantal
UTM-GRID's
van
10
x
10
km²
van
de
periode
van
vóór
1900
en
daarna;
At-1:
aantal
UTM-GRID's
van
10
x
10
km²
van
de
periode
vanaf
1900;
At:
idem
van
1950
t/m heden;
%:
gemiddelde
percentage
afname
van
At
in
de
perioden;
RL:
categorie
Rode
Lijst,
zie
tabel 1.
dankwoord (2003)
Voor
het
opsturen
van
kevers
of
het
doorgeven
van
waarnemingen
bedank
ik
de
volgende
personen:
B.
van
Aartsen,
't
Harde; J.E.F. Asselbergs,
Bergen
op
Zoom; K. van Brakel, Gouda;
Drs.
R. Beenen,
Nieuwegein; C.J.M.
Berger, Hamont-Achel,
België;
Dr.
J.
Bezděk,
Mendel
University
of
Agriculture
and Forestry, Brno,
Tsjechië;
Dr. G.J.C. Buth, Serooskerke;
Drs. J.G.M. Cuppen,
Ede; M.B.P. Drost, Wadenoijen;
A.
van
Dijk,
Amsterdam; L.H. Fliervoet,
Nijmegen; Th. Heijerman, Wageningen;
D. Horsfield,
Nature
Conservancy Council, Edinburgh,
Groot-Brittannië; Drs. J. Huijbregts, Leidschedam;
Drs. R.Ph.
Jansen,
Amsterdam;
K. Jot,
Amsterdam; T. Keukelaar, Wilheminadorp; Dr.
H. Kippenberg, Herzogenaurach,
Duitsland; G.C. Kronenberg, Eindhoven; mevr. J.L. Kuijper-Nannenga,
Arnhem; mevr. M. te Laak, Sas van Gent;
Ir. J.J.J. Langeslag,
Plantenziektenkundige
Dienst,
Wageningen;
C. Langeveld, Hillegom; A. Lutjeboer, Apeldoorn; A.G. Masseur, Hoorn;
F.G.J.M.
van
Nunen,
Vianen; T.M.J. Peeters, Tilburg; P.
Poot,
Maastricht; F.A.H.E. Post, Tilburg;
J.
de
Rond,
Lelystad; W.J.
van Rooyen, Oosthuizen; J.C.P.M. van de Sande, Amsterdam; N.J. Schonewille,
Amsterdam;
H. Silfverberg,
Zoological
Museum,
University
of
Helsinki,
Finland;
D. Sivell,
University
of
York,
Groot-Brittannië; G.J. Slob, Kruiningen;
Drs.
A.
de Smidt,
Harlingen;
Dr. E.A.M. Speijer,
Den
Haag;
Mr. F.C.F.
Sterrenburg,
Den
Haag; A.P.J.A. Teunissen,
Vlijmen; Sj. Tiemersma, Wezep; H.J. Vallenduuk,
Schijndel;
Dr.
P. Verdyck,
Antwerpen,
België; W.J. Veldkamp, Eibergen; D.A. Vestergaard,
Soest; Dr. O.F.J. Vorst, Utrecht; Dr.
A.
Warchałowski,
Instytut
Zoologiczny UWr.,
Wrocław,
Polen; Dr. Ir. J.J. Wieringa, Renkum; J.H.
Woudstra,
Zaandam.
Voor
de
hulp
bij
het
onderzoek
aan
collecties
bedank
ik
Drs.
J.
Fokkema
(Fries
Natuurmuseum,
Leeuwarden);
Y.
Jongema (Wageningen Universiteit); H.C.
Pijpers (Museon,
Den
Haag);
Drs.
J.
Krikken
en
A.
van
Assen
(Nationaal
Natuurhistorisch
Museum
- Naturalis,
Leiden);
F.
van Stuivenberg,
Natuurhistorisch
Museum,
Enschede;
mevr.
Dr. F.N. Dingemans-Bakels,
Natuurhistorisch
Museum,
Maastricht; Ing. B.J.
van
Vondel,
Natuurmuseum,
Rotterdam (NHMR);
Ir. E.H.M.
Bouvy
en T.M.J.
Peeters, Natuurmuseum Brabant,
Tilburg (NNKN); B.J.H.
Brugge
en
W. Hogenes,
Zoölogisch
Museum,
Universiteit
van
Amsterdam,
Amsterdam.
Voor
het
testen
van
de
tabel
bedank
ik
volgende
KNNV-leden
van
afdeling
Amsterdam:
J.
van Arkel,
Amsterdam; M.H.J.
Broer,
Amstelveen;
Dr.
F.
Freese,
Amsterdam;
Drs. V.S.
van
der
Goot,
Amsterdam; W.F.
Klein, Breda; S.C. Langeveld, Hillegom;
Dr.
P. Oosterbroek,
Amsterdam;
mevr. Y.G. Oussoren,
Amsterdam; N.J. Schonewille,
Amsterdam
en
Dr. M.P.
van
Veen, Zeist.
Drs. L.J.J.M. Kemps, Amsterdam dank ik voor alle reflectie.
Mevr.
Dr. S.A.
Ulenberg
van
de
afdeling
Entomologie
van
de
Universiteit
van
Amsterdam
dank
ik
voor
het
beschikbaar
stellen
van
een
tekenmicroscoop
en
werkruimte.
 De
Uyttenboogaart-Eliasen
Stichting verleende financiële steun.
De
Uyttenboogaart-Eliasen
Stichting verleende financiële steun.
 De Nederlandse Entomologische Vereniging voor het
program ORDE (Vorst
& Fokker 1995).
De Nederlandse Entomologische Vereniging voor het
program ORDE (Vorst
& Fokker 1995).
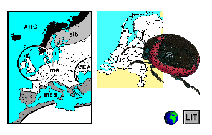
De Nederlandse Goudhaantjes
This site is online since 2003